
Du principe anthropique
à l'homme
En 1859, Charles Darwin publiait son célèbre ouvrage De l’origine des espèces au moyen de la sélection naturelle ou la préservation des races favorisées dans la lutte pour la vie (1), dans lequel il démontrait non seulement que toute forme de vie provient de formes plus anciennes, mais que l’évolution est due à la sélection naturelle, idées simples mais révolutionnaires à cette époque empreinte de créationnisme. Plus tard, les néo-darwiniens expliqueront le mécanisme profond de l’évolution par les mutations génétiques. Dans la même optique, Patrick Delbrouck (2) distingue au sein de la pathologie mentale les maladies « qui correspondent à un fonctionnement chronique, apparemment inadapté à l’environnement, mais persistant dans le temps et dans l’espace au fil des générations, et pour lesquelles se posent à la fois la question du comment, mais aussi celle du pourquoi du maintien de ce “désavantage” par la sélection naturelle. Cette dernière interrogation se rapporte au domaine du darwinisme médical. Son application à la psychiatrie est relativement récente (une vingtaine d’années). Au niveau des psychoses, elle consiste à appréhender l’intérêt évolutif que présenteraient les pathologies productives et/ou déficitaires ; (...) D’emblée, la question à laquelle cette théorie se propose de répondre est radicalement différente de celle posée par la recherche psychopathologique habituelle, même si les deux s’intéressent à “l’origine” des maladies mentales. Le darwinisme se désintéresse du “comment” devient-on psychotique ? pour se recentrer sur le “pourquoi” (...) ? » La question que nous allons nous poser ici est pourquoi l’évolution a-t-elle de tout temps engendré des rétro-adaptations à la vie marine, de plus en plus délicates au fur et à mesure que l’on s’éloigne des poissons dans le phylum, et quel caractère avantageux, au niveau de la survie de l’espèce humaine, peut apporter la persistance de la naissance d’enfants autistes, qui ont d’ailleurs une influence négative sur la fécondité humaine, puisqu’a priori ils ne se reproduisent pas. Bref, ils sont éliminés, mais ils reviennent !
En effet, l’évolution de l’embranchement des vertébrés ne s’est pas faite seulement dans le sens de la conquête programmée des terres émergées, c’est-à-dire du milieu aérien par opposition au milieu aquatique, mais aussi dans le sens d’un retour périodique au milieu marin au prix d’une délicate adaptation à rebours. Prévoyante, la nature semble avoir sauvegardé (dans le sens quasi-informatique du terme), par un retour progressif dans le giron de la mer originelle, une ou plusieurs espèces par classe de vertébrés terrestres au cas où de profondes modifications du milieu aérien risqueraient d’anéantir son œuvre (Tableau 1). Ce pourrait être le cas du cœlacanthe, véritable fossile vivant, dont les ébauches de membres et de poumon évoquent la classe des amphibiens et qui vit encore aujourd’hui au large des Comores. Ce fut le cas de l’ichtyosaure, reptile marin du Secondaire qui présentait de nombreux caractères de convergence avec les dauphins, mais une queue verticale comme celle des poissons (illustration 1). C’est encore le cas, pour la classe des reptiles, des tortues marines (tortue à écailles, tortue luth). C’est le cas du manchot, oiseau nageur plongeant jusqu’à deux cents mètres de profondeur. Le grand pingouin, exterminé au xixe siècle, était lui aussi incapable de voler. C’est le cas bien entendu des mammifères marins : cétacés (baleine, dauphin) et pinnipèdes (otarie, phoque). Les cétacés ne se déplacent pas sur terre, contrairement aux pinnipèdes qui peuvent encore s’y traîner, mais nagent exclusivement grâce aux battements d’une puissante queue horizontale, ce qui les distingue des poissons (queue verticale) malgré d’évidentes convergences morphologiques (illustration 2). Ils ne possèdent pas de cordes vocales, devenues inutiles dans un milieu non aérien.

À l’inverse, un instinct pousse les cétacés, notamment les odontocètes (pourvus de dents et non de fanons : dauphin, orque, globicéphale, cachalot), à s’échouer en masse sur des plages en pente douce dans toutes les parties du monde, où ils meurent étouffés par leur propre poids, comme s’ils tentaient régulièrement de recoloniser le milieu dont ils sont lointainement issus. Ce comportement est à rapprocher du caractère volontiers amphibie des pinnipèdes, des manchots et des tortues. Il peut s’expliquer plus prosaïquement, notamment chez le globicéphale, par l’existence d’un chef de groupe âgé ou malade qui n’a plus la force de nager pour respirer en surface. Croyant trouver le salut en s’échouant, il n’y trouve qu’un bref répit avant d’y étouffer sous son propre poids et surtout y entraîne les autres membres du groupe, conditionnés à le suivre aveuglément, comme un petit suit sa mère étroitement pendant sa première année. La solution est alors de rejeter à la mer tous les membres du groupe, sauf le leader défaillant, car il les y reconduirait inmanquablement. En tout cas, vu la fréquence des échouages de cette espèce, si les terres émergées sont un jour recolonisées par un cétacé, ce sera probablement par un globicéphale mutant.
L’évolution des vertébrés semble donc guidée, on pourrait presque dire “programmée”, derrière le foisonnement apparent des espèces, non seulement par le principe anthropique qui la conduit jusqu’à l’Homme dans une sorte de course à la complexité, mais aussi par une “loi de sauvegarde” qui la pousse à sauvegarder son œuvre à l’apparition de chaque classe de vertébrés par un retour prudent et conservatoire à la vie marine (l’évolution ne serait pas seulement créatrice, elle serait aussi conservatrice), sous forme de véritables “disquettes enregistrées vivantes” (via le génome des espèces préservées) conservées dans le milieu marin. Des caprices plus ou moins dirigés de l’évolution géologique, comme l’isolement d’une population dans un espace insulaire la poussant progressivement à chercher ses ressources alimentaires dans le milieu marin, donc à s’y adapter, pourraient d’ailleurs expliquer cette particularité constante de l’évolution biologique. On aurait ainsi à disposition dans cette “médiathèque marine” :
- la “disquette amphibiens” (cœlacanthe),
- la “disquette reptiles” (tortue marine),
- la “disquette oiseaux” (manchot),
- la “disquette mammifères” (dauphin).
Il va de soi que ces espèces marines sont particulièrement à protéger de la prédation humaine. Mais on remarque aussi que la médiathèque est incomplète et qu’il y manque une disquette, la plus importante à nos yeux et sans doute aux yeux de l’évolution, la “disquette Homme” !
Mais l’évolution, qui ne se dirige pas au hasard mais où le hasard semble plutôt guidé par le principe anthropique, n’a certainement pas oublié l’Homme, sommet de sa création, dans son effort de conservation et de sauvegarde, et nous avons peut-être en fait déjà sous nos yeux les premiers spécimens du futur Homme marin : l’Homo autisticus. Cependant, l’observation des enfants autistes est faite encore de nos jours sous un angle totalement aérien, qui en donne une vision particulièrement négative (3) : leurs troubles relationnels sont très précoces (dès le début de la vie), ils ne se coulent pas dans les bras de leur mère comme les autres nourrissons, ils ne recherchent pas le contact physique (aloneness), ni le contact visuel (ils ne regardent pas en face), ils ne communiquent pas (mutisme, amimie) ou répètent en écho les paroles prononcées devant eux (écholalie), ils ne pleurent pas, ils rejettent tout changement dans leur environnement (sameness), ils apprennent tardivement à marcher, ils préfèrent se balancer d’avant en arrière dans d’interminables mouvements stéréotypés, ils n’ont pas de préférence manuelle (latéralisation défectueuse), ils sont incontinents... et bien sûr inadaptés à la vie terrestre qu’on leur propose. On ajoutera qu’ils n’éprouvent pas d’intérêt pour les visages ni pour la voix humaine, signe précoce aux conséquences relationnelles catastrophiques que nous avons gardé à part, car il a fait l’objet d’une étude en Résonance Magnétique Nucléaire fonctionnelle (4, 5). Celle-ci montre que les autistes utilisent pour la reconnaissance des visages (prosognosie) les aires corticales normalement impliquées dans la reconnaissance des objets neutres et pour la perception de la voix humaine celles impliquées dans la perception des sons ordinaires (au lieu d’activer l’aire spécifique du sillon temporal supérieur).
Mais l’observation de ces enfants peut être faite sous un angle inédit qui se révèle bien plus positif (Tableau 2), si on la considère sous l’angle d’une pré-adaptation à la vie marine, qui pourrait s’expliquer par une fixation de leur propre ontogenèse à un stade précoce de la vie fœtale intra-utérine. Ferenczi ne parlait-il pas de « lambeau d’océan » (Thalassa, 1924) pour désigner le liquide amniotique ? Cet abord paraît conforté par la diminution de la sévérité des troubles autistiques obtenue par Ben-Ari (Marseille) et Lemonnier (Brest) en 2012, grâce à l’utilisation d’un salidiurétique faisant baisser notamment la concentration intracellulaire de chlore dans les neurones (6). Là aussi, il semble bien que le neurone de l’autiste soit en « osmose », c’est le cas de le dire, avec le milieu marin qui est censé l’accueillir. L’augmentation de la natriurèse va faciliter son adaptation au milieu aérien.
Dans cette perspective radicalement nouvelle, l’enfant autiste ne se coule pas dans les bras de sa mère parce qu’il ne se reconnaît pas dans cette mère “aérienne” qu’il vit comme une étrangère et qui, de son côté, le trouve bien étrange : c’est un drame pour la mère comme pour l’enfant, mais l’autiste n’a pas besoin de mère, il a d’abord besoin de mer. Il ne recherche pas le contact physique parce que les membres supérieurs sont plus utiles pour nager que pour étreindre son semblable dans le milieu marin. Il ne recherche pas le contact visuel (à notre façon) parce que l’adaptation hydrodynamique lui imposera plus tard (au cours de l’évolution de son phylum) de passer progressivement d’une vision binoculaire de face à une vision monoculaire et latérale. Il ne parle pas parce que la parole est aérienne et inexploitable sous la surface de l’eau (les plongeurs sous-marins, dont je suis, communiquent par signes).
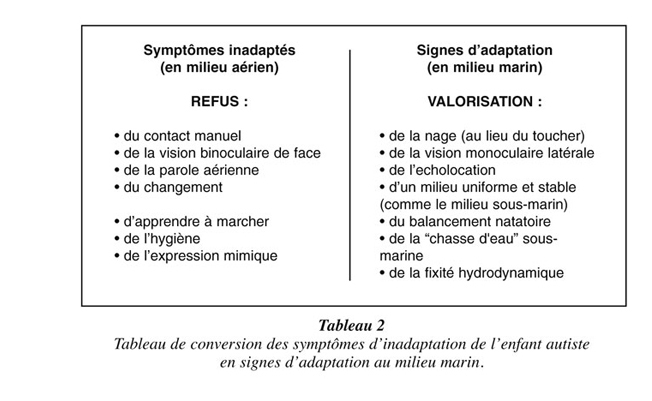
L’écholalie elle-même, lorsqu’elle est présente, préfigurerait les mécanismes d’écholocation des mammifères marins. Les stéréotypies phonétiques, lorsqu’elles existent, sont des consonnes répétitives utilisables sous l’eau (car elles ne nécessitent pas l’ouverture de la bouche comme les voyelles) et rappellent les clics émis en salves par les cétacés à dents comme le dauphin. Les langues primitives à clics des Bochimans (“homme de la brousse”, en afrikaans) et des Hottentots (“bégayeur”, en afrikaans) seraient d’ailleurs un intermédiaire intéressant, autant pour comprendre la communication entre cétacés que pour communiquer avec les autistes (sans passer par la parole articulée). Le claquement de langue utilisé dès l’âge de 6 ans par le jeune aveugle Ben Underwood en Californie ne lui sert pas à communiquer, mais à détecter les obstacles par écholocation, comme les dauphins et pourquoi pas les autistes immergés, et montre que cette adaptation est possible en une seule génération. Ces deux exemples “terrestres” nous montrent que les stéréotypies phonétiques chez l’autiste pourraient avoir une double fonction, comme chez le dauphin : la communication sous l’eau et l’écholocation, gages indispensables d’une adaptation à la vie marine, pour éviter d’être muet et aveugle (sans masque) dans ce milieu a priori hostile.
L’autiste ne pleure pas, parce que les larmes sont inutiles quand les yeux baignent en permanence dans de l’eau salée. Il rejette tout changement autour de lui, parce que le milieu marin profond est un milieu stable et quasiment immuable. C’est d’ailleurs la première impression ressentie par tout plongeur sous-marin : pas de changement de temps ou de saison sous la mer, pas de vent qui se lève et qui plie la végétation, pas de pluie, de neige ou de grêle, pas de tonnerre et d’éclairs, pas de torrent ou de cascade, pas de houle ou de ressac comme en surface, mais des bruits assourdis et des chutes amorties, des tombants où on ne tombe pas (grâce à la poussée d’Archimède), des mouvements ralentis par la pression... L’enfant autiste s’oriente bien dans l’espace, car il est prêt à évoluer dans un milieu à trois dimensions. Il n’est pas motivé pour apprendre à marcher, parce que la marche est inutile en pleine mer. Il aime par contre balancer tout son corps d’avant en arrière, parce que c’est un meilleur moyen pour se déplacer quand le corps est totalement immergé dans l’eau (ex : brasse papillon, nage du dauphin). Les stéréotypies de balancement deviennent ainsi l’ébauche de mouvements natatoires (ondulations du corps qui successivement s’immerge et émerge), subitement adaptés dans l’hypothèse de ce nouveau milieu.
Il est amusant de noter que la prière juive, la âmida, se fait debout et pieds joints, en silence, et un balancement du tronc d’avant en arrière apparaît naturellement pour rythmer les mots égrenés à voix basse. Il s’agit ici d’un autisme volontaire, d’un repli de l’homme sur sa relation avec Dieu. Qu’il soit volontaire ou involontaire, on voit que les effets de l’autisme sont les mêmes et s’expriment dans un syndrome commun, fait de silence et de balancement, pieds soudés.
Le balancement natatoire sagittal des cétacés est le mouvement le plus stéréotypé de la lasse des mammifères avec la respiration, à laquelle il est d’ailleurs lié (ils doivent se soulever régulièrement hors de l’eau pour respirer).
Le balancement sagittal de l’autiste n’est donc pas une simple stéréotypie psychomotrice, c’est la préfiguration d’un mouvement de survie essentiel dans le milieu marin (nage + respiration). Autre point commun, contrairement à la marche qui est un mouvement discontinu (on ne tombe pas en s’arrêtant et on repart facilement), le balancement de l’autiste est un mouvement continu qui peut se prolonger pendant des heures car, comme l’avait démontré Louis de Corlieu (l’inventeur génial des palmes de natation) dès 1907, la discontinuité de l’impulsion diminue l’efficacité de la propulsion dans l’eau et un mouvement continu est alors préférable, non seulement pour rester en sustentation et pour respirer en surface, mais aussi pour économiser l’énergie nécessaire à l’avancement dans le milieu.
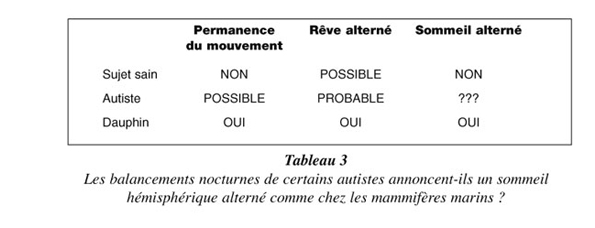
Certains autistes ne dorment pas ou paraissent ne pas dormir et se balancent ainsi toute la nuit, assis sur leur lit, rappelant la permanence des mouvements natatoires que doivent entretenir les mammifères marins pendant leur sommeil, sous peine de se noyer ! Le dauphin ne dort en effet que d’un hémisphère à la fois (chaque hémisphère dort alternativement pendant 20 mn) pour continuer à venir respirer en surface (d’où la permanence des mouvements natatoires) et parce que sa respiration est volontaire et non automatique comme chez l’Homme (pour ne pas respirer sous l’eau ou dans une vague). Il serait intéressant d’enregistrer les tracés électroencéphalographiques de l’autiste insomniaque, pour y déceler les signes d’un sommeil persistant mais asymétrique comme chez ce mammifère marin. Une anomalie à rapprocher de cette nécessité vitale, qui d’après Michel Jouvet se produit chez deux tiers des sujets sains, est la dissociation des visages et des messages dans le sommeil paradoxal, c’est-à-dire qu’on reconnaît un acteur du rêve sans comprendre ce qu’il dit, ou inversement on entend ce qu’il dit sans le reconnaître. Cette anomalie ne peut s’expliquer que parce que les visages sont liés à l’hémisphère droit du cerveau (reconnaissance spatiale), tandis que les messages sont liés à l’hémisphère gauche (centres du langage), ce qui revient à dire que le cerveau rêve tantôt avec l’hémisphère droit, tantôt avec l’hémisphère gauche. Cette disposition naturelle, ou plutôt cette prédisposition, vraisemblablement commune à l’autiste et à la plupart des sujets sains rappelle à un niveau voisin la structure du sommeil orthodoxe chez le dauphin, qui dort en alternance tantôt avec l’hémisphère droit, tantôt avec l’hémisphère gauche, pour contrôler sa respiration avec l’hémisphère resté éveillé. Du rêve alterné au sommeil alterné, il n’y a qu’un pas que la nature a pu franchir avec certains autistes comme elle l’a fait avec les mammifères marins (Tableau 3)...
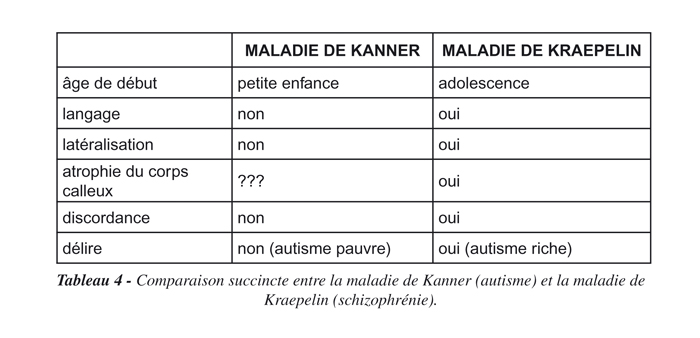
Les troubles présentés par les enfants autistes sont très précoces, mais c’est parce que dans notre hypothèse ils doivent s’adapter dès la naissance au milieu marin (dans lequel ils sont censés être nés). Ils sont ambidextres parce que leurs mains leur serviront à nager et non à saisir, et que la nage est un mouvement symétrique. Ils n’ont pas de cerveau dominant comme les enfants normaux, car de toute façon ils n’utilisent pas le langage et parce que s’ils doivent un jour nager en dormant, l’hémisphère qui reste alternativement en éveil devra être polyvalent. On peut d’ailleurs se demander si, comme cela vient d’être démontré chez le schizophrène (7), une atrophie du corps calleux qui relie les hémisphères cérébraux entre eux ne s’installe pas parallèlement à la maladie. Elle serait responsable (ou la conséquence) de l’absence de latéralisation, mais ne provoquerait pas de discordance comme lorsque la maladie (regardée comme un continuum entre l’autisme et la schizophrénie) se déclare à l’adolescence, car le sujet est alors latéralisé et ses hémisphères spécialisés. La schizophrénie pourrait être considérée, de façon quasi-étymologique, comme une scission fonctionnelle entre des hémisphères spécialisés, d’où la discordance de la pensée observée à cet âge et révélée par le langage. Ce qui nous permet de dresser un tableau comparatif succinct des deux maladies, ou de la même maladie se déclenchant à des âges et à des stades de développement différents (Tableau 4) :
Ils sont incontinents, mais les cétacés ne vont pas non plus aux toilettes puisque, dans leur milieu, la “chasse d’eau” est tirée en permanence ! Ils n’éprouvent pas d’intérêt pour l’expression des visages, car les contraintes hydrodynamiques imposeront la fixité des traits au cours de leur évolution régressive à l’intérieur du milieu marin, qui font que même le sourire permanent du dauphin Tursiops est un “objet neutre” dépourvu de toute signification affective (donc sans intérêt). Les plongeurs sous-marins eux-mêmes n’accordent aucun intérêt aux visages de leurs congénères déformés ou écrasés par la pression, et donc dépourvus de mimique significative. On voit donc que l’inadaptation de l’enfant autiste est plus éducative que physiologique, pour peu qu’on substitue un milieu marin au milieu aérien dans lequel on veut l’éduquer. Ils adorent d’ailleurs aller à la piscine ou à la mer, où ils ne craignent pas de rester en immersion et gardent longtemps les réflexes archaïques du “bébé nageur” : bouche fermée et yeux ouverts. Le risque de noyade est très faible (sous surveillance) et on peut dire avec certitude, si l’on en croit leur excitation, qu’ils y sont plus heureux qu’en milieu aérien. Enfin, leur intelligence paraît normale, si on ne les teste pas avec les tests verbaux et de performance habituels.
L’enfant du silence pourrait donc bien en fait être adapté au monde du silence. Un silence relatif bien entendu, car si les poissons sont muets, les cétacés ne le sont pas (chant des baleines, sifflements et cliquetis des dauphins). On mesure aussitôt les conséquences médico-éducatives qui découlent de cette hypothèse. En fait, ces implications sont déjà mises en pratique empiriquement par le biologiste israélien Roni Zilberg et par l’équipe de Sophie Donio, soutenue par le psychiatre israélien Daniel Meir, au Dolphin Reef d’Eilat, au bord de la mer Rouge (8). Le projet baptisé “Supportive experience with the aid of dolphins” a démarré en 1991, un an après l’ouverture des portes du Dolphin Reef, s’inspirant d’un projet déjà existant mené depuis 1989 par le Docteur David Nathanson à Key Largo en Floride. Ce programme y prenait en charge en mai 2002, lorsque je m’y suis rendu, des enfants et des adolescents autistes au contact de treize dauphins de l’espèce Tursiops truncatus (photo) vivant dans un espace naturel d’un hectare, profond de dix-huit mètres et ceinturé (mais non fermé : il existe une porte, ce qui fait l’originalité de cet enclos) par un filet dont les mailles laissent passer les poissons nécessaires à leur nutrition. Le but n’est pas de faire de ces enfants des apprentis dauphins ou des Hommes marins, mais d’améliorer la communication entre l’enfant et ses parents ou son thérapeute, via cet animal dans lequel il s’identifie volontiers et qui vient naturellement vers lui et son “dolphin trainer” dès les premiers contacts.

L’équipement de l’enfant est habituellement sommaire : combinaison isotherme qui, associée à l’exceptionnelle salinité de la mer Rouge (40 g/l), assure la flottabilité du patient. Mais, pour des prises en charge plus prolongées, rien n’empêche d’imaginer l’équipement traditionnel du “snorkeling” : un masque pour améliorer la vision, un tuba pour aller chercher l’air au-dessus de la tête (comme les évents du dauphin) et même des palmes soudées pour parfaire le mimétisme avec l’animal (la queue horizontale) et mieux utiliser les stéréotypies de balancement du jeune autiste. L’important est surtout de remotiver l’enfant à la communication (voire d’utiliser ses capacités verbales) et de lui apporter le plus tôt possible le déclic nécessaire pour relancer son développement psychomoteur dans un milieu approprié à sa pathologie, et lui éviter de s’enfermer dans une “psychose de milieu” qui va obérer ses acquis et stériliser à jamais son intelligence. Toutes les tentatives de rééducation médicale de ces enfants devraient en fait être initiées dans ce type de structure, où la mer (à 25°C en moyenne) est le support et le dauphin l’éducateur naturel de l’enfant autiste, sous la surveillance du “dolphin trainer”. Dans cette perspective rééducative réalisée dans un milieu approprié respectant les spécificités évolutives de l’enfant autiste (au lieu de s’y opposer vainement) et même ses convergences évolutives avec le cétacé, sous stricte surveillance médicale spécialisée et sans espoir affiché de guérison, mais seulement de réadaptation comportementale dans son milieu par destination, on peut alors parler véritablement de “delphinothérapie” pour l’enfant autiste. Notre projet serait de transposer cette expérience en France, grâce au plateau technique du Marineland d’Antibes, même si les sept dauphins qui y vivent le sont dans une stricte captivité et entraînés avant tout pour le spectacle. La ville d’Antibes est d’ailleurs jumelée avec celle d’Eilat et, autre symbole fort, Franck Esposito le champion d’Europe 2002 du deux cents mètres nage papillon, ou nage du dauphin, est un nageur antibois. La thérapie serait proposée sous forme de séances de flottage ou de nage en présence de l’animal, de caressage, grâce à la médiation de l’entraîneur du dauphin, éventuellement de nourrissage, voire de remorquage (en s’accrochant à l’aileron dorsal) dans les cas les plus favorables, le tout étant accompagné d’une écoute liquidienne des bruits émis par les cétacés, soit directe pendant la séance, soit indirecte par casque et microphone immergé. En fait, on peut dire que les autistes sont mal traités dans le monde entier, faute de moyens adaptés mis à leur disposition, excepté par les psychiatres Daniel Meir et David Nathanson (déjà cités), mais aussi l’éthologiste Lorenzo von Fersen au delphinarium de Nuremberg (Allemagne, 1998) et le psychologue Richard Griffiœn au delphinarium d’Harderwijk (Pays-Bas, 2001), où je me suis rendu en août 2002. J’aimerais en France être capable à mon tour, dans l’un des deux delphinariums existants, de leur apporter ce dont ils ont réellement besoin pour mieux vivre dans ce monde aérien qui les comprend si mal, par incapacité ou manque de volonté à s’identifier à leur si étrange problématique (la plus étrange sans doute de toute la pathologie mentale).
Le choix du dauphin nous a paru supérieur, même s’il est plus difficile à mettre en œuvre, à celui d’animaux vivant en milieu aérien (comme le cheval) pour trois raisons :
- le milieu marin recrée les conditions de la vie intra-utérine (le bassin et l’eau de mer, qui rappellent le bassin pelvien et l’eau de la mère, c’est-à-dire le liquide amniotique) et j’ai observé que l’enfant doit être attiré dans l’eau dans sa langue maternelle (par exemple en russe, au lieu de l’anglais ou de l’hébreu, si l’enfant est un immigré russe israélien),
- le dauphin est un mammifère amical qui manifeste un désir spontané de contact avec l’espèce humaine sans attente de nourriture (les dauphins du Dolphin Reef sont nourris à heures fixes 4 fois par jour, sans rapport conditionnant avec la thérapie, et sortent du bassin pour chasser les espèces qu’ils prisent particulièrement dès qu’ils les ont repérées avec leur sonar),
- on peut aider l’enfant autiste à retrouver les racines du désir de communiquer en lui faisant écouter les bruits sous-marins émis par les cétacés et captés à l’aide de microphones placés dans le bassin, qui réactivent en lui l’écoute “in utero” porteuse du désir originel de communiquer (écoute liquidienne inspirée de la méthode Tomatis).
On peut se demander pourquoi aucune tentative n’a été réalisée avec les singes anthropoïdes (chimpanzé). C’est parce que le grand singe a en commun avec l’Homme la vision binoculaire de face, qui fait fuir le regard de l’autiste et gêne tant la communication avec ses propres congénères, fussent-ils ses parents.
Pour terminer, sur un plan éthique, notons qu’en cas de typage génétique futur de la maladie, il conviendra de proscrire tout mode de reconnaissance in utero et surtout de traitement prophylactique par sélection prénatale des embryons (eugénisme scientifique), quelque soit le coût de la prise en charge du handicap. En effet, si l’évolution retient avec constance ce type de constitution pathologique dans la biodiversité humaine (4 à 5 autistes sur 10 000 enfants en France comme dans d’autres pays), c’est peut-être pour préserver tout le génome humain en cas de catastrophe écologique majeure à la surface des terres émergées, qu’elle soit d’origine naturelle (ex : chute d’une météorite géante) ou provoquée par l’Homme lui-même : guerre nucléaire, disparition de la couche d’ozone, submersion des continents due à la fonte des glaces (effet de serre), etc. En effet, si l’on estime à 6 milliards d’individus la population mondiale, dont la moitié vivant sur une bande côtière de 30 km de large, et à 40 % les jeunes de moins de 15 ans, cela fait tout de même 480 000 enfants autistes vivant près du littoral. Même si un faible pourcentage de ces enfants parvenait grâce à sa pré-adaptation à survivre d’abord dans un milieu estuaire ou lagunaire proche de la (nouvelle) côte, puis en pleine mer, ce faible contingent suffirait à repeupler le nouvel écosystème et, par la reproduction au sein du groupe, à affirmer les traits autistiques pré-existants et ainsi à consolider l’adaptation au milieu. Le fait que les généticiens soupçonnent les gènes Hox (qui sont impliqués aussi dans l’évolution néoténique du préchimpanzé à l’australopithèque, puis à l’Homme) A1 et B1 sur les chromosomes 7 et 17 comme pourvoyeurs de l’autisme, semble d’ailleurs indiquer qu’il s’agit plus d’une mutation évolutive que d’une maladie au sens propre du terme. Il s’agit sans doute d’une nouvelle “néoténie”, c’est-à-dire d’un ralentissement de la vitesse de développement du jeune autiste l’empêchant de se métamorphoser en sujet aérien et le retenant à un stade aquatique du développement fœtal.
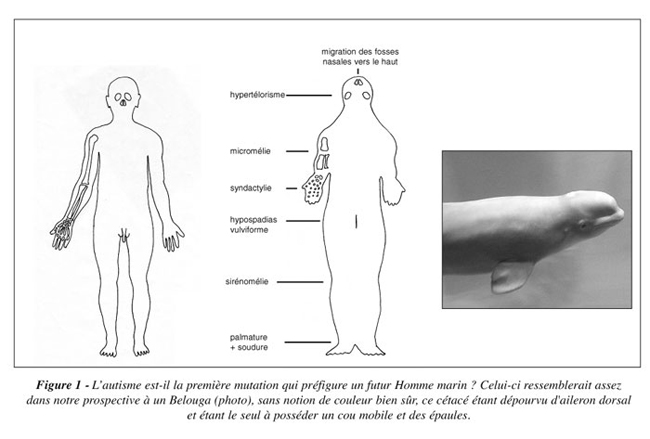
On notera enfin que des malformations congénitales d’ordre physique, cette fois, peuvent aussi prédisposer à une adaptation à la vie marine et, conjuguées à l’autisme, pourraient parfaire cette adaptation (Figure 1) :
- l’hypertélorisme, écartement anormal des yeux qui préfigure le glissement latéral des yeux vers l’arrière, par adaptation fusiforme hydrodynamique (torpedo-like shape) ;
- le pterygium colli, replis cutanés bilatéraux tendus de la région mastoïdienne à la région acromiale de chaque côté du cou (adaptation fusiforme et portance) ;
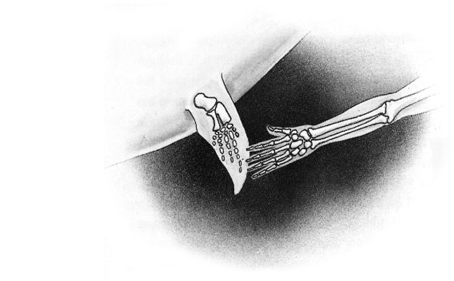
- la micromélie segmentaire symétrique, qui arrête le développement en longueur des membres supérieurs (illustration);
- la palmature des doigts (paume est voisin de palme), ou syndactylie, et des orteils ;
- l’hypospadias vulviforme, qui retire le pénis en dedans comme chez les cétacés et le met à l’abri sous une pseudo-fente vulvaire, pour éviter toute aspérité génératrice de turbulences ;
- la sirénomélie, qui soude les membres inférieurs entre eux du pubis à la plante des pieds (1/70 000 naissances).
Il n’est pas interdit de penser, conformément à la théorie saltatoire de l’évolution (Gould), que la conjonction de cette inadaptation comportementale à la vie aérienne que représente l’autisme, de ces différentes malformations congénitales et de nouvelles conditions environnementales puissent conduire à une adaptation beaucoup plus rapide, peut-être aussi brutale que l’évènement déclenchant, qu’on ne le pense ordinairement : à un véritable “saut évolutif” en quelque sorte.
Le tropisme réciproque et les convergences évolutives de l’autiste et du dauphin placeront peut-être un jour les delphinidés au centre du monde humain. Delphes n’était-elle pas considérée autrefois comme le nombril de la Terre (omphalos) ? D’autant plus que les cas de sauvetage de nageurs imprudents et de marins naufragés ramenés sur la plage par les dauphins sont légion depuis l’Antiquité, depuis le sauvetage du poète Arion jeté par-dessus bord au large de Corinthe (tout près de Delphes) et le sauvetage de Jonas par la baleine qui le “vomit” ensuite sur le rivage (Jon 2). Ils pourraient d’ailleurs jouer un rôle déterminant en secourant certains autistes au début de leur rétro-adaptation à la vie marine, dans les circonstances dramatiques que l’on imagine.
L’idée n’est pas plus folle, elle est plus poétique en tout cas, que celle de Laurent Mottron (L’autisme : une autre intelligence, 2004) qui soutient, en faisant allusion aux capacités spéciales des autistes savants (ex : calcul de calendrier), que l’autisme est destiné à devenir un avantage culturel en milieu simplement aérien : « il existe un important faisceau de résultats indiquant que la prédisposition à l’autisme et, dans une mesure qui reste à déterminer, l’autisme lui-même peut représenter un avantage adaptatif. (...) l’ensemble des domaines de traitement de l’information pour lesquelles les personnes autistes sont supérieures aux non-autistes au niveau perceptif et mnésique (...) en constitue l’argument le plus solide. » Cet argument en forme de “canular”, pour reprendre le terme discourtois utilisé par le québecois Mottron à l’égard de mes propres travaux, sert de couverture scientifique à des intérêts colossaux au service de la méthode TEACCH, qui prétend (ré)éduquer les enfants autistes à grand renfort de personnel spécialisé (non psychiatrique) et d’institutions largement subventionnées par le budget public. Il existe donc un véritable “marché” de la souffrance des enfants autistes et de leurs parents que se disputent aujourd’hui la méthode TEACCH (Schopler), la psychanalyse qui est la méthode la plus ancienne (Bettelheim) et la delphinothérapie qui est la plus récente (Nathanson). C’est en partie la raison de l’échec de mon projet de prise en charge de jeunes autistes au Marineland d’Antibes. Donc, je résume : 1° pour Mottron, l’autisme est un avantage adaptatif au monde moderne ; pour moi, l’autisme est un avantage adaptatif au monde marin / 2° pour Bettelheim, l’autiste est une “forteresse vide” ; pour moi, l’autiste serait plutôt un “sous-marin en cale sèche”. Faites votre choix !